

The Role of Nrf2 in Cellular Defense Against Environmental Stressors
The Nrf2 pathway is one of the important genomic regulators protecting the cell from environmental stressors by managing the antioxidant and detoxification processes within most cells.1 Like most complex genomic regulatory systems, Nrf2
(nuclear factor erythroid 2-related factor 2) is sequestered in the cytoplasm until it is able to translocate into the nucleus, where it forms a heterodimer with MAF (another transcription factor) and binds to the promoter region of genes containing
an antioxidant response element (ARE) sequence. Nrf2 regulates important genes such as heme-oxygenase 1(HO-1), glutathione S-transferase (GST), glutathione reductase (GR), glutamate-cysteine ligase subunits (GCLc and GCLm), NAD(P)H quinone oxidoreductase-1
(NQO1), and thioredoxins (TrxR1), among others.2,3 Nrf2 stays sequestered in the cytoplasm by its interaction with the Keap1 protein complex (Keap1-Kelch-like ECH-associated protein 1). When an oxidative stressor or electrophile interacts
with the Keap1-Nrf2 complex, it allows for the release and subsequent nuclear accumulation of Nrf2 through a complex process of ubiquitination-inhibition, thus triggering an anti-stress response by the cell.4 In some circumstances,
Nrf2-activation can mitigate some of the effects of the NF-κB inflammatory signaling pathway.5,6
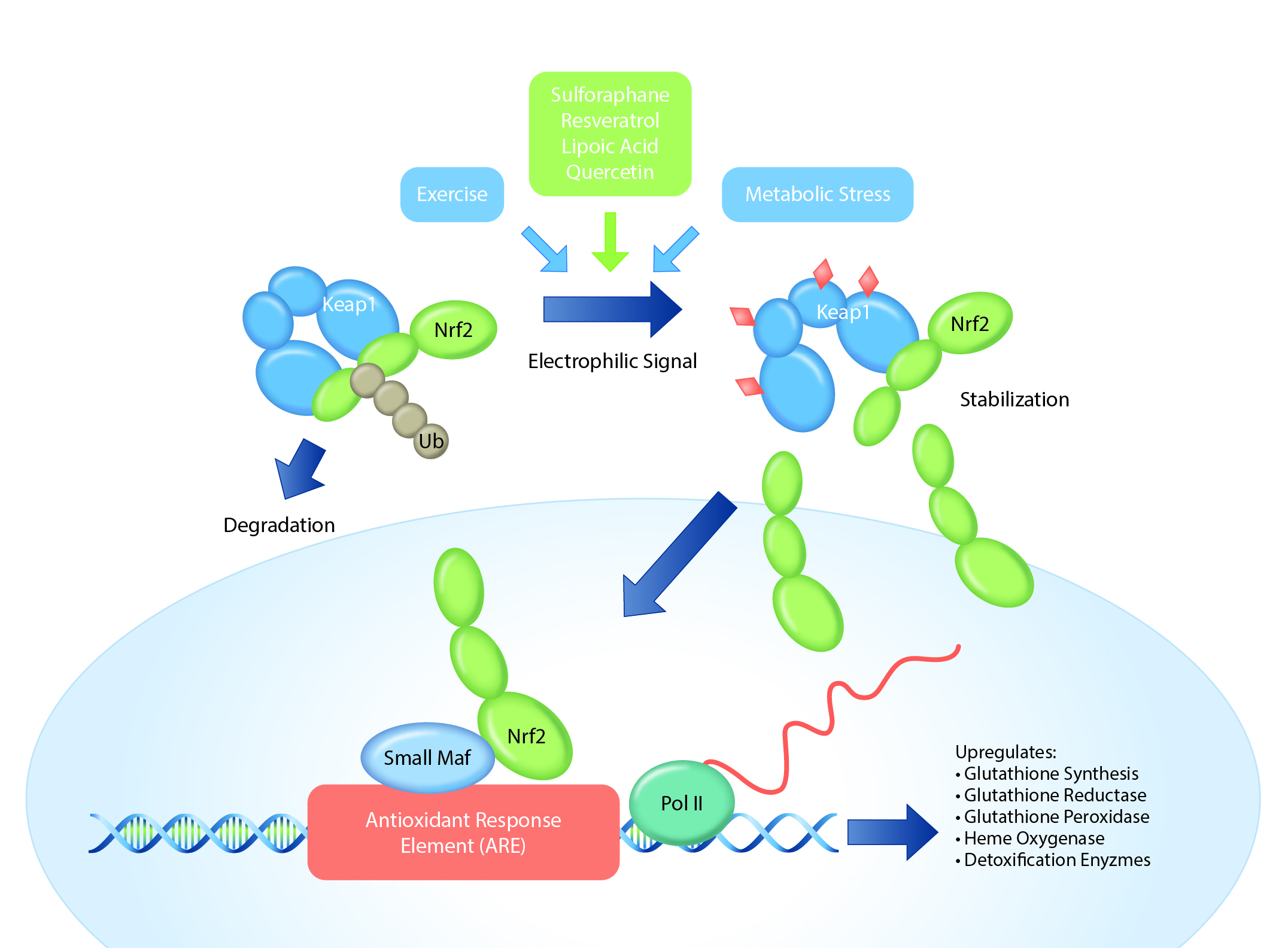
The Keap1–Nrf2 system. Under normal conditions, Nrf2 is constantly ubiquitinated through Keap1 and degraded in the proteasome. When exposed to electrophiles (i.e., certain antioxidants) or oxidative stress, Keap1 is inactivated. Stabilized Nrf2 accumulates in the nucleus and activates transcription of many genes that produce antioxidant proteins. Ub, ubiquitin. Image adapted from: Mitsuishi Y, Motohashi H, Yamamoto M. The Keap1-Nrf2 system in cancers: stress response and anabolic metabolism. Front Oncol. 2012 Dec 26;2:200.

Natural Compounds That Modulate Nrf2 Pathways
There has been a wealth of research looking at agents (naturally occurring and synthetic) that can upregulate this global antioxidant and detoxification system. Sulforaphane (a glucosinolate compound found in some cruciferous vegetables like broccoli) is one of the most commonly cited natural Nrf2 activators, though there are many others shown to have similar effects on Nrf2-induced enzymatic activities and metabolites. There are well over 100 different nutrients, phytonutrients, and related dietary supplement ingredients thought to function as antioxidants or detoxification enhancing agents, at least in part, because of their ability to up-regulate Nrf2 activation. A small portion of these have been confirmed to alter such responses when consumed at reasonable oral doses in human or animals and are listed below:7
- Cruciferous glucosinolates (broccoli, Brussels sprouts, etc.)8,9,10
- Sulforaphane (concentrated)11,12
- Curcuminoids (turmeric)13,14
- Quercetin15,16
- Resveratrol17,18
- Phenethyl isothiocyanate (PEITC, from watercress)19,20
- Diallyl sulfide (garlic)21,22
- Lipoic acid23,24
- N-acetyl cysteine (NAC)25,26

Additional Nrf2-Modulating Ingredients
The list of natural compounds shown to modulate Nrf2 pathways or ARE-containing genes is too numerous to list in this text, though common ingredients for which the reader might be familiar include all those listed above and also baicalein (Scutellaria baicalensis), catechins (tea, cocoa), genistein (soy), carnosol and carnosic acid (Rosmarinus officinalis), andrographolide (Andrographis paniculata), ginsenoside Rb1 (Panax ginseng), schisandrin B (Schisandra chinensis), quercetin and related flavonoids, indole-3-carbinol (I3C) and diindolylmethane (DIM), hydroxytyrosol (Olea europaea - olive), Pterostilbene, Xanthohumol (Humulus lupulus), berberine, [6]-gingerol (Zingiber officinale), vitamin D, and even exercise (though the Nrf2-activation response decreases with aging).26,27,28,29
The Challenge of Specific Dosage Recommendations for Nrf2 Activation
Content from this blog has been adapted from Supplementing Dietary Nutrients: A Guide for Healthcare Professionals.

Thomas G. Guilliams, PhD (Tom) earned his doctorate in molecular immunology from the Medical College of Wisconsin in Milwaukee. For the past two decades, he has spent his time investigating the mechanisms and actions of lifestyle and nutrient-based therapies, and is an expert in the therapeutic uses of dietary supplements. Tom serves as an adjunct assistant professor at the University of Wisconsin School of Pharmacy and was the VP of Science for Ortho Molecular Products for 24 years (he now serves them as a consultant). Since 2014 he has been writing a series of teaching manuals (Road Maps) that outline and evaluate the evidence for the principles and protocols that are fundamental to the functional and integrative medical community. He is the founder and director of the Point Institute, an independent research and publishing organization that facilitates the distribution of his many publications. A frequent guest-speaker, Dr. Guilliams provides training to a variety of health care disciplines in the use of lifestyle and natural medicines. He lives in the woods outside of Stevens Point, Wisconsin with his wife and children.

